|
Черноголовый хохотун (Larus ichthyaetus Pallas, 1773) Great Black-headed Gull (англ.)
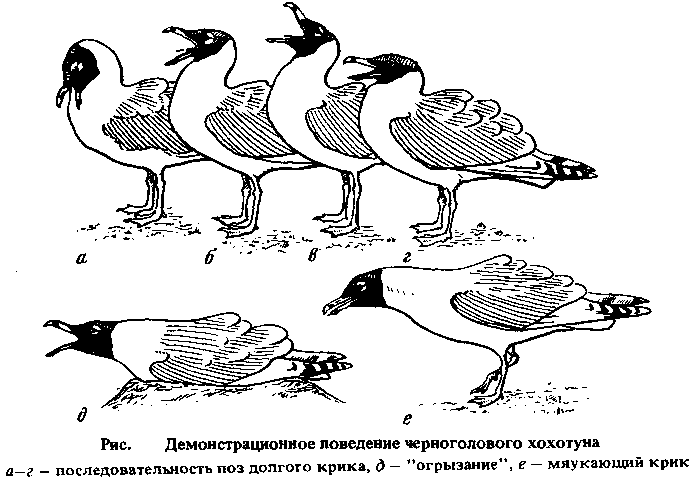
Статус Статус. Гнездящийся мигрирующий, на Каспии частично оседлый вид. Общая характеристика и полевые признаки. Одна из самых крупных и красивых чаек нашей страны и мировой фауны, по размерам близка к бургомистру и большой морской чайке. От других крупных чаек хорошо отличается бархатисто-черной головой, от темноголовых чаек — очень большими размерами. По рисунку первостепенных маховых хохотун сходен с реликтовой чайкой, от которой легко отличается размерами и трехцветным клювом (оранжево-желтое основание, красный кончик и черная перевязь у вершины). Летает, медленно взмахивая крыльями, может парить. Хорошо плавает, на воде сидит высоко, не ныряет. Очень осторожен, вне колонии молчалив. Голос очень низкий: в колонии и ее окрестностях можно слышать низкое "гау" или "ау", долгий крик напоминает глухой рев. Вопреки названию и некоторым литературным данным и в отличие от серебристой чайки-хохотуньи черноголовый хохотун не хохочет — если не считать хохотом короткий тревожный крик "ха-га-га". Монотипический вид.Описание. Строение и размеры. Размеры (мм) и масса тела (г): Длина крыла: самцов (п = 17) - 485-517 (среднее 503); самок (n = 16) - 462-493 (среднее 472). Длина клюва: самцов (п = 18) - 59,1-70,3 (среднее 64,2); самок (п = 15) - 47,5-65,3 (среднее 56,7). Длина цевки: самцов (п = 18) - 77,6-88,6 (среднее 81,4); самок (п = 16) - 68,4-84,9 (среднее 73,6). Масса тела: самцов ( п = 8) - 1130-2000 (среднее 1600); самок (п = 8) - 960-1500 (среднее 1220). Окраска. Самец и самка в брачном наряде. Голова бархатисто-черная, причем черный цвет спереди слегка захватывает верх шеи, а сзади весь затылок. Над и под глазами узкие белые полоски. Шея, грудь, брюхо, бока, надхвостье, подхвостье и рулевые белые. Спина, плечевые и верхние кроющие крыла бедные сизовато-серые, задние плечевые с белыми вершинами. Первостепенные маховые белые, кроме серых трех последних. II (первое видимое) первостепенное с черным на большом протяжении наружным опахалом и черным предвершинным пятном, иногда не доходящим до внутреннего края опахала. На III расположено почти такое же пятно, не доходящее до внутреннего края опахала, и, кроме того, маленькое черное пятнышко почти у самой вершины. Черные предвершинные пятна (полосы) есть на IV—VII. Второстепенные маховые серые с белыми вершинами. Клюв оранжево-желтый с предвершинной черной перевязью, конец клюва красный. Лапы зеленовато-желтые. Радужина бурая, края век ярко-красные.Самец и самка в зимнем наряде . Лоб, темя, затылок, уздечка темно-бурые, с густыми белыми пестринами (могут быть и белыми с бурыми пестринами), кроющие уха однотонные темно-бурые; горло и подбородок белые. Остальное оперение, как в брачном нарядеПуховой наряд . Птенец целиком серебристо-белый или кремово-белый, нередко сверху с охристым оттенком. Изредка на спине и крыльях неясные светло-серые пятна. Лапы черновато-серые, клюв черный с желтоватой вершиной.Гнездовой наряд . Лоб, темя, затылок покрыты серо-бурыми перьями с серыми каемками, в результате чего голова кажется сероватой с неясными более темными пестринами. Такие же, но более крупные перья есть на шее и спине. Плечевые и кроющие крыла, кроме больших, серовато-бурые с охристыми каймами; большие кроющие крыла серые. Серовато-бурый цвет перьев может быть разной интенсивности у разных особей, нередко с коричневатым оттенком. Подбородок, горло, бока, брюхо, надхвостье белые, грудь белая с бурыми пестринами. Первостепенные маховые темно-бурые, начиная с III на внутреннем опахале в средней части пера появляется светлое поле. Второстепенные маховые бурые с белыми вершинами. Рулевые белые с темно-бурой верхней третью. Клюв буроватый у основания с почти черной вершинной частью. Лапы серовато-бурые, радужина темно-бурая, края век черные.Первый зимний наряд . Лоб, подбородок, горло, грудь, бока, низ шеи, брюхо, надхвостье белые. Темя белое с бурыми пестринами, которые становятся густыми на затылке и задней части шеи. Перед глазом и на кроющих уха бурое пятно. Перья спины и плечевые становятся сероватыми с более темными бурыми центрами. Крылья и рулевые сохраняются от гнездового наряда.Первый летний наряд . Как первый зимний, но на голове появляются темные буроватые перья.Второй зимний наряд . Лоб, темя, горло, подбородок, надхвостье и нижняя части туловища белые. Затылок белый с темно-бурыми пестринами, шея белая с отдельными крупными темно-серыми пестринами. Перед глазом и на кроющих уха нечеткое темное пятно. Спина серая с буроватым оттенком и отдельными бурыми пестринами. Первостепенные маховые черные с белыми основаниями и белыми полями на внутренних опахалах; на II и III первостепенном обычно появляются небольшие белые предвершин ные пятна. Рулевые белые с широкой черной предвершинной полосой, иногда отсутсвующей на паре боковых перьев.Второй летний наряд . Голова черно-бурная с белыми пестринами, густота которых индивидуально варьирует. Остальное оперение, как в предыдущем наряде.Третий зимний и третий летний наряды , как окончательные, но на первостепенных маховых (особенно на II и III) черный рисунок развит больше, а на рулевых у част особей сохраняются остатки черных предвершинных пятен.Линька. Во время частичной линьки в первый зимний наряд в сентябре—декабре сменяется оперение головы и туловища, оперение крыла и рулевые остаются от гнездового наряда. Последующие линьки бывают дважды в году: частичная предбрачная, во время которой не меняются рулевые и оперение крыла, и полная послебрачная. Предбрачная линька взрослых бывает, очевидно, между январем и мартом, послебрачная между июнем и декабрем (возможно, январем). Полные и частичные линьки в промежуточные наряды проходят, по-видимому, в несколько более ранние сроки. Распространение. Гнездовой ареал. Область гнездования простирается от северных районов Крыма до котловины Больших озер в Монголии и оз. Кукунор (по некоторым данным до среднего течения р. Хуанхэ) и от оз. Чаны до южной части Каспия. Сплошного ареала вид не образует, для него характерны отдельные гнездовые поселения, нередко удаленные на несколько сот километров друг от друга. Одни поселения существуют в течение десятков лет, другие возникают на короткое время при улучшении гидрологического режима водоемов. На севере Крыма хохотуны гнездятся на Сиваше и Лебяжьих островах (Каркинитский залив Черного моря). На Сиваше гнездование известно с начала 30-х годов: в 1932—1934 гг. на островах у Чонгарского моста и, по крайней мере, с конца сороковых годов по настоящее время на о-ве Китай. На Лебяжьих островах гнездование регистрируется с 1947 г. По материалам ЦКАН, в 1977 г. существовала колония в Кирпильском лимане Приморско-Ахтырского района Краснодарского края. В 1983 г. отмечено гнездование на Кривой косе в Донецкой области. С начала 1960-х годов хохотун стал регулярно отмечаться на гнездовье на озерах и водохранилищах долины Маныча: на оз. Маныч-Гудило, Пролетарском и Чограйском водохранилищах, на оз. Казинка и Меклетинских озерах Калмыкии, а также на расположенных неподалеку рыбхозах. В то же время на Сарпинских озерах, где хохотун гнездился ранее, он ныне не обнаружен. На Каспийском море в настоящее время гнездится лишь на севере Каспия: на о-ве Жемчужном, в дельте Урала и на островах северо-восточного побережья Каспия от Гогольской косы до п-ова Дурнаев. Крайне нерегулярно гнездится в дельте Волги. В 1952 г. отмечено гнездование на Тюленьих (Мангистауских) островах, современное состояние этого поселения неизвестно. На юге Каспия, где прежде гнездование отмечалось в заливе Кирова (Азербайджанское побережье), и, по-видимому, в Красноводском заливе, на Челекене и о-ве Огурчииском ныне не гнездится. Гнездовое поселение в заливе Кара-Богаз-Гол, существование в 1979—1980 гг., исчезло после перекрытия пролива, соединявшего залив с морем. В Казахстане постоянно гнездится на озерах Тенгиз (Целиноградская обл.), Алаколь, Балхаш. В 1948 г. отмечена колония в 3 тыс. особей на о-ве Комсомольский в Аральском море. Последующая судьба этой колонии, как и колоний в устье р. Сырдарьи и по восточному побережью Аральского моря, неизвестна. Судя по исчезновению колонии на о-ве Барсакельмес, поселения хохотунов на Аральском море сейчас испытывают период депрессии. Подобное наблюдается на оз. Зайсан, где эти птицы ныне перестали гнездиться, хотя прежде, по-видимому, были многочисленны. В 1962 г. найдена колония на оз. Жалаулы в Павлодарской области. Севернее известно лишь одно постоянное поселение хохотунов — на оз. Чаны в Новосибирской области.Весьма вероятно (хотя и не доказано) гнездование черноголовых хохотунов на озерах в низовьях Тургая, а также на оз. Карасор Карагандинской обл.. В Киргизии гнездование этих птиц возможно на оз. Иссык-Куль у мыса Кара-Булун. Среди возникающих на короткое время поселений отмечены гнездовья на оз. Соркуль восточнее пос. Джаныбек (Уральская обл.) в 1968—1971 гг. и 1981 г, на оз. Жарколь (Кустанайская обл.) в 1966-1968 гг., на оз. Таволжанном (юг Тюменской обл.) в 1959-1961 гг.. На основании опросных данных высказано предположение о гнездовании хохотунов на оз. Джулукуль на юго-востоке Алтая в 1962—1964 гг.; в 1977 г. хохотуны здесь не гнездились. 24. V 1985 г. одиночное гнездо черноголового хохотуна найдено в колонии серебристых чаек и речных крачек на о-ве Таволжанском Запорожского водохранилища. Вопреки первоначальным предположениям, хохотун не гнездится на оз. Маркаколь, на Памире и, по-видимому, на Камыш-Самарских озерах, хотя в последнем случае возможны нерегулярные поселения. Вне территории бывшего СССР достоверно гнездится на озерах Хара-Ус-Нур, Толбо-Нур и Убсу-Нур, очень вероятно гнездование на оз. Хяргас-Нуре. Гнездование указывалось также для озер Урэг-Нура, Ачит-Нура и Бон-Цаган-Нура, однако неясно, основываются ли эти сведения на находках гнезд или же, как в случае Ачит-Нура, на встречах птиц в гнездовом наряде (что для хохотуна не может служить доказательством гнездования). Вопреки прежним представлениям, на оз. Хубсугул (Косо-гол) хохотун не гнездится. Для Китая отмечается гнездование на оз. Кукунор, на озерах истока р. Хуанхэ и в среднем течении этой реки (Внутренняя Монголия) также без указания на находки гнезд. Что касается оз. Кукунор, то гнездование хохотунов здесь можно считать доказанным. Мнение же о гнездовании этих птиц на озерах верховий р. Хуанхэ, высказанное впервые Н.М. Пржевальским (1888), до сих пор находками гнезд не подтверждено. В отношении динамики ареала хохотуна можно, очевидно, пока говорить лишь о перераспределении колоний внутри области гнездования, а не о каком-либо изменении ее границ. Исключение составляет лишь юг Каспия, где эти птицы перестали гнездиться, скорее всего, вследствие изменения уровня Каспия. Однако то, что хохотун оказался способным гнездиться на водохранилищах, показывает потенциальную возможность расширения ареала в будущем, — о чем, в частности, говорит недавняя попытка гнездования на Запорожском водохранилище. В последние годы участились случаи встреч чаек этого вида на Куйбышевском водохранилище. Зимовки. Основной район зимовки — Каспийское море (главным образом его южная часть), откуда получено 12 (40%) из 30 зимних (декабрь—февраль) возвратов хохотунов, окольцованных в Казахстане; 23,3% зимних возвратов поступило с внутренних водоемов Ирана (в основном из озерно-болотной местности Заболь), 13,3% — с водоемов юга Средней Азии (Хаузханское водохранилище, Каракумский канал, Айдарские разливы, Амударья в Сурхандарьинской обл.), 6,7% — из Индии и по 3,3% — с внутренних водоемов Калмыкии и Азербайджана, Эфиопии (оз. Тана), Кувейта и южного побережья Аральского моря. Анализ данных, поступивших в ЦКАН, показывает, что с юга Каспия (включая разливы р. Артек) получено 34,4% из 32 зимних возвратов хохотунов, окольцованных на севере Каспия, из северных и северо-западных районов Каспия — 21,9%, с внутренних водоемов Волгоградской и Саратовской областей — 15,6%, из внутренних районов Ирана и Эфиопии (оз. Тана) — по 6,3%, и по 3,1% с внутренних водоемов Азребайджана, северного Ирака, побережья Красного моря в ЙАР, Омана (Салала) и из г. Батуми. От хохотунов, окольцованных на водоемах Маныча, получено по два зимних возврата из Астраханской обл. и южных районов Каспия, по одному — с внутренних водоемов Ростовской обл. и Сирии, побережья Израиля, Персидского залива, из Эфиопии (оз. Тана) и Индии. Основное место зимовок хохотунов северного Причерноморья, видимо, тоже Каспийское.В целом с учетом данных кольцевания и визуальных наблюдений в область зимовки этого вида следует включить южную часть Средиземного моря (а не только египетское побережье), Красное море, внутренние водоемы Эфиопии, морские побережья от Адена и Персидского залива до Бирмы, внутренние водоемы Сирии, Ирака, Ирана и Пакистана, незамерзающие участки Каспийского и изредка Аральского морей, внутренние водоемы Волгоградской, Саратовской и Ростовской областей, юга Средней Азии и Казахстана. Зимующие особи отмечены на оз. Иссык-Куль. В Черном море у берегов Крыма зимовки носят, по-видимому, нерегулярный характер.Миграции. Покидает места зимовок в конце февраля — начале марта, хотя отдельные особи появляются в окрестностях гнездовий, расположенных по соседству с районами зимовок, уже в феврале: 11—16.11 на Лебяжьих островах, 11.11 на о-ве Барсакельмес. Пролет взрослых особей завершается в сжатые сроки: к концу марта — началу апреля на оз. Чаны, однако, первые птицы появляются лишь в середине — конце апреля. Неполовозрелые птицы начинают миграцию одновременно со взрослыми, но их пролет сильно растянут и завершается в конце апреля — первой половине мая. Пролет части казахстанской популяции хохотунов, по-видимому, может идти через Центральную Азию — оз. Лобнор и озера Тибета. На это как будто указывает выраженный пролет в северном направлении через Джунгарские ворота в конце марта — апреле. Первые хохотуны на оз. Лобнор отмечены 19.11, на оз. Кукунор — в начале — середине марта.После подъема молодых на крыло (июнь — июль) хохотуны начинают широко кочевать, нередко залетая на сотни километров за пределы области гнездования вида. Преобладающее направление кочевок казахстанских хохотунов в июле—августе западное и северо-восточное; направленный пролет отмечается уже в августе, в последующие месяцы он приобретает все более выраженное западное и юго-западное направление (некоторые особи долетают до Азовского моря). В большинстве районов Казахстана пролет завершается в октябре. Сходная картина наблюдается у хохотунов с оз. Чаны. У птиц с Лебяжьих островов направление осеннего пролета восточное, в сторону Каспия. Большинство молодых хохотунов, окольцованных на севере Каспия, держатся в районе рождения, часть кочуют довольно далеко в западном, юго-западном и северном (вплоть до Татарстана) направлениях. По данным ЦКАН, хохотуны с водохранилищ системы Маныча до сентября держатся главным образом в Ростовской обл. (66% возвратов), часть откочевывает на север до Волгоградской и Саратовской областей, другие появляются западнее — на побережье Азовского моря, юго-западнее и южнее — в Краснодарском и Ставропольском краях; на побережье Каспия отмечаются лишь единичные особи. В октябре — ноябре птицы распределены по тому же обширному району, но более равномерно; встречи на севере Каспия становятся более частыми. Широкие послегнездовые кочевки особей из разных поселений, захватывающие почти всю область гнездования, наряду с общими местами зимовки позволяют предположить, что все черноголовые хохотуны Казахстана представляют единую популяцию, внутри которой не исключена возможность перемещения птиц из одних гнездовых колоний в другие. По тем же причинам к этой популяции должны относиться птицы с Каспия и оз. Чаны. Не исключено, что все хохотуны, гнездящиеся на территории бывшего СССР, представляют собой единую популяцию. В период гнездовых кочевок взрослые и молодые птицы нередко в массе появляются на богатых рыбой озерах, где отсутствовали на гнездовье: оз. Маркаколь на Южном Алтае, оз. Зоркуль и Яшилькуль на Памире, оз. Малый Чан в Новосибирской области и др.. Как в сезон гнездования, так и вне его отмечены многочисленные залеты черноголовых хохотунов за пределы гнездового ареала и области зимовок. К дальним залетам можно отнести встречи птиц в Англии, Бельгии, Нидерландах, Дании, Швеции, Польше, Италии, Греции, Румынии, на о-ве Мадейра, на Западной Украине, в восточной части Латвии, в Томской обл., Борзинском районе Читинской обл. (ЦКАН), Гонконге, на оз. Виктория в Уганде. Численность. На севере Причерноморья гнездится до 290 пар, на водохранилищах системы Маныча — около 1800 пар, возможно несколько больше. На о-ве Жемчужном в разные годы гнездится от 3500 до 10 тыс. пар, а всего на севере Каспия — около 10 тыс. пар или несколько больше. На Кара-Богаз-Голе в 1979 г. гнездилось 628 пар. На оз. Тенгиз в последние годы гнездилось 343—875 пар, на оз. Алаколь — 2250 —3700 пар, на оз. Балхаш — 2500—3000 пар, на оз. Чаны — 72—190 пар. Всего в пределах бывшего СССР гнездится, видимо, 20—25 тыс. пар. Общая тенденция изменения численности не ясна в связи со значительными несинхронными колебаниями ее в разных частях ареала. Постоянное увеличение численности отмечено для северных районов Причерноморья; на оз. Алаколь после роста в 1970-х годах она, видимо, стабилизировалась; на оз. Тенгиз в целом снижается. Численность вне СССР неизвестна.Местообитание. В сезон гнездования — крупные соленые и солоновато-водные озера, морские мелководья и реже водохранилища в степной полупустынной и пустынной зонах. Для гнездования необходимы изолированные от наземных хищников острова, для кормежки, — по-видимому, мелководья с непостоянным уровнем воды либо иные участки, где хохотуну легко добывать рыбу не ныряя или собирать ее в большом количестве полуснулой. Во внегнездовое время — морские побережья, долины рек, озера, водохранилища, пруды рыбхозов.Размножение. К размножению хохотуны приступают, по-видимому, на четвертое лето, отдельные особи, возможно, на год раньше. Черноголовый хохотун — облигатно-колониальный вид; моногам. Характерны резкие колебания численности птиц в колониях по годам, связанные с перераспределением гнездящихся пар в пределах одного озера или ближайших озер и, возможно, с негнездованием части особей. Не исключено и перераспределение пар между поселениями в разных частях ареала в зависимости от изменения гидрологического режима водоемов.Черноголовые хохотуны гнездятся только на островах (как на останцах, так и плоских, косах), причем нередко удаленных на несколько километров от берега. Под гнездовья выбираются лишенные растительности возвышенные участки. Хохотуны практически всегда гнездятся вместе с серебристыми чайками, которые поселяются по краю их компактных колоний или в непосредственной близости от них. По соседству с колониями хохотунов могут гнездиться и другие чайковые птицы. На территории колонии можно наблюдать различные формы демонстрационного поведения, которое у хохотунов имеет ряд своеобразных черт. Характерная особенность хохотунов во время выполнения различных демонстраций — взъерошивание перьев спины, плечевых и третьестепенных маховых, в ряде случаев — разворачивание веером хвоста. Долгий крик хохутуна достаточно своеобразен. Его можно разделить на четыре фазы; в первых двух издается протяжный ревущий крик. Общая продолжительность ритуала 3—5 с. Как и у других чаек, долгий крик хохотуна полифункционален. Основная агрессивная демонстрация — "огрызание", или ложное клевание, — выпад раскрытым клювом в сторону противника . К сфере брачного поведения относятся "кашляние" или "квохтанье", и мяукающий крик. В позе мяукающего крика хохотун издает протяжное "охх-ггооо" (или "ухх-ггууу" при закрытом клюве). Данная демонстрация отмечается во время брачных церемоний, при смене партнеров на гнезде и кормлении птенцов. "Квохтание" похоже на аналогичную демонстрацию серебристой чайки.
Как и у других чаек, у хохотунов в период формирования пар и реже во время насиживания можно наблюдать кормление самки отрыгнутой самцом пищей; выпрашивание корма сопровождается вскидыванием клюва. Прямая поза и отворачивание головы ритуализованы слабо, "распластанная" поза и "клевание травы" отсутствуют. Величина колоний хохотунов варьирует от нескольких десятков до нескольких сот пар. Реже отмечаются колонии в несколько пар, гнездование отдельными парами бывает исключительно редко и всегда в колонии других видов чайковых. Гнезда в колониях располагаются пространственно разделенными группами — субколониями, в которые входит от нескольких до нескольких сот гнезд, по пространственной конфигурации различаются плотные многорядные, плотные линейные и разреженные линейные колонии. Плотность гнездования очень велика. Среднее расстояние между центрами ближайших соседних гнезд в колониях севера Причерноморья составляет 1,0 м (n = 200), индекс плотности гнездования 0,32. Со сходной плотностью хохотуны гнездятся и в других частях ареала: так, в заливе Кара-Богаз-Гол среднее минимальное расстояние между краями гнезд составляло в разных субколониях от 0,39 до 2,13 м, в большинстве случаев 0,4—0,9 м, на оз. Чаны среднее расстояние между центрами ближайших соседних гнезд (n = 72) было 0,92 м. В качестве материала для гнезда используются сухие водоросли, сухие стебли травянистой растительности, отдельные перья. В начале насиживания в гнезде может не быть выстилки; достройка гнезда продолжается весь период насиживания. Наружные стенки гнезд в период насиживания всегда запачканы пометом. Размеры гнезд (мм) : на оз. Тен-газ диаметр гнезда 500—700, диаметр лотка около 260, глубина лотка 70—80; на оз. Жарколь диаметр гнезда (n = 18) 290—582, средний 445, диаметр лотка 250—354, средний 299, глубина лотка 56—83, средняя 72; на Лебяжьих островах (n=16) диаметр гнезда 350—560, средний 449, диаметр лотка 220-280, средний 253, глубина лотка 65-110, средняя 88. Откладка яиц в Крыму начинается в конце марта — начале апреля, в Казахстане на оз. Жарколь 13—19. IV, на оз. Тенгиз 16. IV, на о-ве Барсакельмес 20. IV. В случае гибели гнезд могут быть повторные кладки. Полная кладка содержит 1—3, чаще всего 3 яйца; на Лебяжьих островах очень редко отмечается 4 яйца. Окраска яиц, по И. А. Долгушину (1962), в целом гораздо светлее, чем у серебристой чайки. По беловатому с кремовым или желтоватым налетом, реже серовато-оливковому фону разбросаны серовато-фиолетовые глубокие и буроватые или черноватые поверхностные пятна. Размеры яиц (мм): с Лебяжьих островов (n = 200) 70,5—88,3 х 483—56,7, в среднем 77,3 х 53,8; с Сиваша (n = 226) 72,0—90,7 х 49,2—58,4, в среднем 78,6 х 54,1; с оз. Тенгиз (n = 50) 72—86 х 51—57, в среднем 78 х 54,5; с оз. Жарколь (n = 36) 70—83 х 40—59, в среднем 77,9 х 53,8, масса свежих яиц (n = 3) 120—127 г, в среднем 123 г; с Аральского моря (n = 10) 66,7—82 х 52—55,2 в среднем 77,9 х 54,1. Насиживают оба партнера. На оз. Жарколь смена происходила несколько раз в день, самки сидели без смены по 3-7 ч и более, самцы — по 1—4 ч. На Сиваше, по наблюдениям за 4 гнездами в конце периода насиживания, смены происходили 2-3 раза в сутки, причем промежутки между ними достигали 17 ч 42 мин (возможно, из-за удаленности мест кормежки); ночью насиживают и самки и самцы. Насиживание с первого яйца, первый птенец вылупляется через 25—29 сут, в среднем через 27 сут (n = 13, Лебяжьи острова). Между появлением трещины на яйце и вылуплением проходит 1-5 сут, чаще всего 3 или 2 сут (50% и 29% соответственно; n=78), между появлением отверстия в скорлупе и вылуплением — главным образом (75%) 1 сут. Между вылуплением первого и последнего птенца в гнезде с кладкой из трех яиц проходит 1—6 сут, чаще всего 2 сут (48,5% случаев), реже 1 сут (21,2%) и 3 сут (15%). После вылупления птенцов родители не уносят скорлупу, ее либо затаптывают, либо выталкивают на край гнезда. На Лебяжьих островах первые птенцы появляются 11—19.IV (птенцы повторных кладок 17—23.V1), на Сиваше 17.IV, на оз. Тенгиз с 6.V по 1—5.VI, на оз. Жарколь 15.V, в заливе Кара-Богаз-Гол 30.IIV. Вылупление птенцов в каждой отдельной колонии протекает обычно в достаточно сжатые сроки — от 11 до 21 дня. Масса тела однодневных птенцов на оз. Тенгиз составляет 40—80 г, на оз. Жарколь 82—90 г. На 4-6-е сутки после вылупления старшего птенца выводки оставляют колонию и кочуют по берегу в стайке с другими выводками. В эти же сроки (на 4—5-е сутки) формируется взаимное опознавание родителей и птенцов. Во время тревоги птенцы в первые дни жизни затаиваются в гнезде или непосредственной близости от него, а после оставления колонии сбиваются в плотный табунок. Поднимаются на крыло в возрасте около 45 дней. Первая попытка кормления птенца отмечена через 3 ч после вылупления. По наблюдениям на Сиваше, в первый день жизни птенцов кормят 11 раз в сутки (данные по 1 гнезду), в возрасте 1—2 сут — 27—30 раз (2 гнезда), 5—6-суточных — 15—27 раз (2 гнезда), 13—17-суточных — 6—16 раз (2 гнезда). Родители кормят птенцов круглосуточно, чаще всего с 9 до 21 ч (76% всех кормежек). Самцы кормят в среднем в 1,4 раза чаще самок (n = 132). Родители обогревают (в жару затеняют) птенцов до 11—13-суточного возраста, при этом смена партнеров происходит 4—7 раз в сутки, в среднем (n = 7) 4,7 раза. После становления индивидуального опознавания родители агрессивно относятся к чужим птенцам своего вида, нередко забивая их насмерть. В разных колониях количество погибших по этой причине птенцов варьирует от 1% до 21,3% и более от числа вылупившихся. Отмечены случаи забивания родителями насмерть своих птенцов, чаще всего младших в выводке. Стратегия защиты потомства у хохотуна носит в основном пассивный характер на основе "эффекта плотности" (отпугивающее воздействие плотной массы сидящих птиц) и направлена против неспециализированных пернатых хищников типа серебристой чайки. Черноголовые хохотуны значительно менее агрессивны, чем более мелкие серебристые чайки, однако в редких случаях способны пикировать на человека, находящегося в колонии. Эмбриональная и постэмбриональная смертность варьирует в разных поселениях. На оз. Жарколь в 1967 г. через месяц после начала вылупления осталось в живых 47,5% птенцов от числа отложенных яиц, на Сиваше в 1973 г. — 23,2%, на оз. Тенгиз в 1976-1978 гг. — 7,4-21,8%, в заливе Кара-Богаз-Гол в 1979 г. — 45-50%; процент неоплодотворенных яиц в различных колониях варьирует от 1,4% до 33,3%. На Сиваше из 102 погибших птенцов 45,1% съедены серебристыми чайками, 3,0% раздавлены родителями в гнезде, 17,6% оставлены родителями и погибли от голода, 3,0% убиты взрослыми хохотунами, 31,3% исчезли (очевидно, также съедены серебристыми чайками). Здесь же разорено серебристыми чайками от 21,9% до 32,8% кладок. На оз. Тенгиз в 1976-1978 гг. в условиях дефицита корма в качестве основной причины гибели птенцов указывалось уничтожение их взрослыми хохотунами, в том числе каннибализм; не исключено, однако, что в последнем случае оказалась неучтенной хищническая деятельность серебристых чаек. По данным кольцевания, смертность молодых в первый год жизни составляет 80%, в последующие примерно по 50% ежегодно. Максимально известный возраст черноголового хохотуна 16 лет 3,5 месяца. Суточная активность, поведение. В сезон гнездования активность, по-видимому, круглосуточная, хотя в светлое время суток хохотуны несколько более активны, чем в темноте. На оз. Жарколь и на Сиваше неоднократно наблюдали ночные смены партнеров на гнезде, кормления птенцов, прилет птиц в колонию, их отлет; на оз. Жарколь отмечена ночная кормежка. В сезон гнездования черноголовые хохотуны очень общественные птицы, однако в период кочевок и зимовки они обычно встречаются одиночно или небольшими группами. Большие стаи ( до нескольких сот особей) изредка могут образовывать отдыхающие птицы на пролете и зимовках. Трофически в большей степени связан с водоемами, чем с сушей. В большинстве случаев в пищевом рационе хохотуна преобладает рыба, мелкие грызуны, насекомые, птенцы водоплавающих птиц имеют подчиненное значение. На поедание хохотунами наряду с рыбой большого количества саранчи и жуков во время массового появления этих насекомых указывали Е.П. Спангенберг и ГА. Фейгин (1936). В 94 погадках, собранных в 1973 г. у гнезд хохотунов на о-ве Китай (Сиваш), рыба встречена в 84,0%, грызуны — 8,5%, птицы — 1,1%, наземные насекомые — 2,1%, речной рак — 1,1%, падаль — 9,6%, остатки растений — 3,2%. В 62 погадках, найденных в 1974 г. у гнезд этого вида на Лебяжьих островах, рыба составляла 93,6% встреч, череп степного хорька — 1,6%, пух птенца — 1,6%, наземные насекомые — 3,2%, растения — 24,2%.Однако есть данные, что животные суши могут играть большую роль в питании хохотуна в некоторых районах в случае ухудшения кормовых качеств водоемов. Анализ 521 погадки на оз. Жарколь за 1966—1968 гг. и содержимого 9 желудков показал, что млекопитающие (из которых преобладает водяная полевка) встречены в 81,8% погадок, рыбы (в основном карась) — в 43,9%, птицы и их яйца — в 15,8%, насекомые (главным образом, жуки-водолюбы) — в 28,6%. Не исключено, однако, что погадки черноголовых хохотунов оказались немного "разбавленными" погадками серебристых чаек-хохотуний, гнездившихся поблизости. Во всяком случае, в 9 желудках хохотунов была встречена только рыба (14 карасей и 2 окуня) и 3 жука. Что касается нередко цитируемой работы И.С. Солдатенко(1956), из которой следует, что остатки наземных животных встречаются в 91,44% погадок хохотунов с о-ва Китай, а рыбы — только примерно в 20%, то почти наверняка за погадки хохотунов были в большинстве случаев приняты погадки серебристых чаек. За это говорит большое сходство в спектре питания этих двух видов, обнаруженное И.С. Солдатенко, большое количество (634) погадок, собранное всего от 51 пары птиц за малый период времени, и сбор погадок в период, когда выводки хохотунов уже покинули колонию и перемещались по острову. Наблюдений за кормовым поведением хохотуна мало. По-видимому, он способен улетать кормиться за десятки километров. По Д. И. Чекменеву, здоровую рыбу в глубокой воде хохотун поймать не может — разве что в местах ее больших скоплений или на высыхающих водоемах, где подвижность рыбы ограничена. Нередко хохотун добывает рыбу из сетей или поедает отбросы промысла. На озерах кормится как в одиночку, так и стаями. Отмечена ловля насекомых (жуков) в воздухе в сумерках.В норме хохотун в отличие от серебристой чайки крайне редко поедает птенцов своего вида, но в отдельных случаях в условиях недостатка корма каннибализм, по-видимому, может принимать значительные размеры. Враги, неблагоприятные факторы. Серьезное воздействие на колонии черноголового хохотуна оказывает серебристая чайка-хохотунья, особенно при усилении фактора беспокойства. Яйца и птенцов чаек этого вида в отдельных случаях могут уничтожать лисицы, барсуки и кабаны. Отмечалось затопление колоний во время нагонных ветров и штормов. Из-за своих крупных размеров и рыбоядности хохотун до сих пор еще нередко попадает под выстрел, на что указывает достаточно высокий процент возврата колец. Хозяйственное значение, охрана. Несмотря на рыбоядность, отрицательного хозяйственного значения в силу малочисленности не имеет. Вид занесен в Красную книгу РФ.
|