|
Озерная чайка (Larus ridibundus Linnaeus, 1766) Black-headed Gull (англ.) С и н. (рус.): речная чайка, обыкновенная чайка.
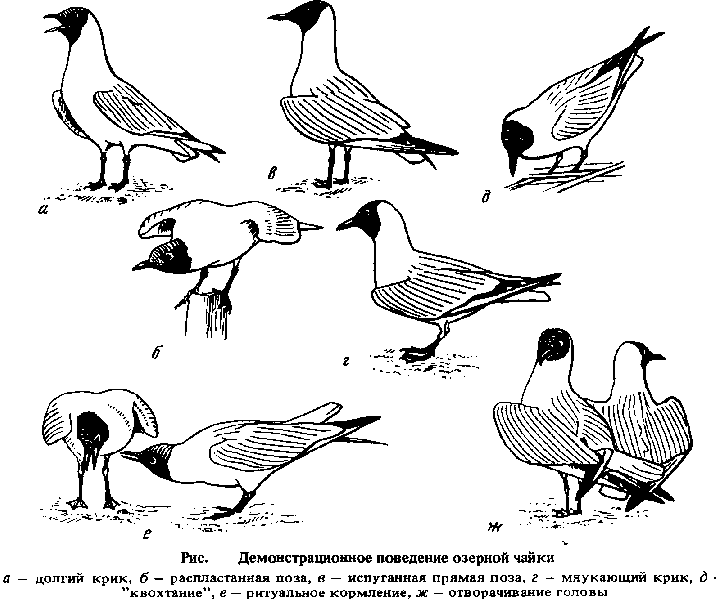
Статус Статус. Гнездящийся, мигрирующий вид, в южной части ареала — зимующий. Общая характеристика и полевые признаки. Чайка средних размеров (немного меньше черноголовой чайки и морского голубка, примерно на 25% меньше сизой чайки). В брачном наряде имеет темно-коричневую голову (издали выглядит черной), граница коричневой окраски на шее у стоящей птицы косая - идет от затылка к горлу (у малой и черноголовой чаек головы в брачном наряде черные, включая затылок, в связи с чем граница черного цвета почти горизонтальная). Концы крыльев черные (у черноголовой и малой чайки белые), нижняя сторона крыльев относительно светлая (у малой чайки темно-серая). Хорошим отличительным признаком во всех нарядах служит клинообразно расширяющийся в дистальной части ярко-белый передний край крыла, образованный первостепенными маховыми и их кроющими. В зимнем наряде по окраске очень похожа на морского голубка, но отличается от него относительно более короткими шеей и клювом, более высоким лбом. Молодые птицы по характерному рисунку верхней стороны крыльев хорошо отличимы от молодых птиц других видов чайковых (у похожего морского голубка верхняя сторона более светлая). В Центральной Азии следует при определении учесть сходство с несколько более крупными буроголовой чайкой и реликтовой чайкой. Подвиды. Монотипический вид. Описание. Строение и размеры (Московск. обл., оз. Киево). Длина крыла (среднее): самцы (n = 66) 319,1; самки (n = 91) 303,1. Длина клюва (среднее): самцы (n = 65) 36,2; самки (n = 90) 33,0. Масса: самцы (n = 23) 293; самки (n = 37) 257. Окраска. Самец и самка в брачном наряде. Голова до затылка, подбородок и горло темно-коричневые. Глаза сверху и снизу окаймлены узкой белой полоской. Передняя и средняя часть спины и верхняя часть крыла серые. В дистальной части крыла клинообразное белое пятно, расширяющееся к концу крыла. Концы II—VII первостепенных маховых черные, на конце VIII субтерминальное черно-серое пятно. Остальная часть оперения — шея, нижняя часть тела, хвост, надхвостье — белые. Клюв, края век, ноги — темно-красные, радужина бурая. Взрослые самец и самка в зимнем наряде. Как в брачном наряде, однако голова белая. Глаз спереди окаймлен черным. Под глазом (иногда и за ним) и в области ушных перьев черно-серые пятна. Пятна у глаз (менее выражены пятна у ушных отверстий) могут быть связаны темной полосой, проходящей поверх головы. Клюв светло-красный с темным концом, ноги светло-красные. Пуховой наряд. Охристо-бурый (варьирует в довольно широких пределах) с крупными черно-бурыми пятнами различной формы. Клюв грязно-мясного цвета с темным концом, ноги тоже грязно-мясные. Гнездовой наряд. Голова светло-бурая, более светлая в передней и нижней части. На ней можно различить более темное пятно перед глазом, полосу под глазом и пятно в области уха, более светлое пятно за глазом. Задняя сторона шеи и передняя часть спины серовато-бурая с желтоватыми концами перьев. Задняя часть спины серая с переходом в белое в области надхвостья. Хвост белый с темно-бурой предвершинной полосой шириной 15—25 мм, за который следует узкая желтоватая каемка на самом конце перьев (на крайней паре рулевых предвершинное пятно может отсутствовать или быть небольшим). Плечевые перья бурые с более светлым желтоватым окаймлением и серые. Второстепенные маховые с темными концами. Первостепенные маховые черновато-бурые с белым (II-V) и серым (VI-VII) клинообразным рисунком, который на VIII—XI маховых становится расплывчатым. Верхние кроющие крыла белые, серые и разных оттенков бурого и черновато-бурого цвета. Нижние кроющие серые, на переднем краю крыла белые. Нижняя часть тела белая со светло-буроватой расплывчатой полосой поперек груди. Клюв грязно-желтый с темным концом, ноги грязно-желтые. Первый зимний наряд. Как зимний наряд взрослых, но хвост и крылья, а иногда и плечевые сохранены от гнездового наряда. Первый летний наряд. Как первый зимний наряд, но голова у большинства птиц бурая с белыми пестринками, особенно на лбу, горло часто почти белое. Ювенильный рисунок на крыльях и хвосте стал более светлым в результате износа. Во втором зимнем наряде птицы иногда отличаются от взрослых, по темным отметинам на крыльях (в основном на кроющих наружных первостепенных маховых). Птицы в брачном наряде с белыми пестринками на голове, по-видимому, двухлетние. Более подробные описания приводятся в литературе. Линька. Послебрачная линька у взрослых полная (конец мая—ноябрь), предбрачная —частичная, сменяется оперение головы и туловища (январь—март, иногда затягивается до мая). Линька в первый зимний наряд частичная: сменяется оперение головы, туловища, часть кроющих крыла (июль—декабрь). Предбрачная после первой зимовки частичная: сменяется оперение головы, туловища, часть кроющих крыла, иногда центральная пара рулевых (январь—май). Послебрачная годовалых: как послебрачная у взрослых, но на 3—4 недели раньше. Распространение. Гнездовой ареал. Сильное расширение ареала началось с XIX в., но особенно проявилось в XX в., что связано предположительно с потеплением климата, улучшением охраны во многих странах и освоением чайками новых антропогенных источников кормов. Граница современного ареала на севере вне территории бывшего СССР влючает о-в Ньюфаундленд (первое гнездование в 1977 г.), юго-запад Гренландии (с 1969 г.), Исландию, Фарерские острова, Британские острова, проходит по северу Скандинавского полуострова и Финляндии. На территории бывшего СССР северная граница ареала включает Онежское озеро и проходит около Архангельска, включает верховья Вычегды, проходит на Урале около 60° с.ш., около 67° с.ш. на Оби, 65° с.ш. на Енисее, 65° с.ш. на притоке Вилюя — Мархе, 68° с.ш. на Колыме. Гнездится на Камчатке и к северу от полуострова в Корякском нагорье, а также на о-ве Карагинском. Отсутствуют данные о гнездовании на обширных территориях на западном берегу Охотского моря, но гнездится на его южном побережье, например в нижнем течении Амура. По Г.П. Дементьеву (1951), гнездится на Сахалине, но более поздние авторы этого не подтверждают. Южная граница гнездового ареала на западе включает отдельные гнездовья в Испании, на юге Франции, на островах Сардиния и Сицилия, долину р. По в Италии, север Югославии и Болгарии, отдельные гнездовья в центральной и восточной частях Турции. На территории бывшего СССР граница ареала проходит по северному побережью Черного моря (отсутствует в Крыму), включает Закавказские республики, огибает с севера Каспий, проходит по дельте Волги, через Камыш-Самарские озера, Актюбинск, спускается к Аральскому морю, включает его северную часть, Сырдарью, равнинные районы Южного и Восточного Казахстана, озера Иссык-Куль и Сон-Кёль в Киргизии. Гнездовые поселения отмечены также на оз. Айдар-куле. Вне бывшего СССР южная граница далее проходит по Монголии, где озерная чайка гнездится на озерах и реках западной, северо-западной и центральной частей страны, на оз. Буйр-Нур и Восточной Монголии, включает северо-восток Китая и советское Приморье. Зимовки. Популяции, гнездящиеся к северу и востоку от январской изотермы -2,5° С, являются мигрантами, гнездящиеся на Британских островах и в бассейне Средиземного моря оседлы, у населяющих промежуточные области молодые птицы в большинстве случаев мигрируют, а у взрослых в направлении с востока на запад прослеживается возрастающая тенденция к оседлому образу жизни. В массовом количестве зимуют к югу и западу от изотермы 0 °С. Таким образом, в область зимовки озерной чайки входят почти все страны Европы, где это позволяет ледовая обстановка зимой, Средиземноморье, побережья Черного, Каспийского морей, а также морей Индийского и Тихого океанов, омывающих южные и восточные (к югу от 45° с.ш.) берега Азиатского материка. За последние десятилетия чайка стала зимовать на побережье Северной Америки от о-ва Ньюфаундленд до Нью-Йорка, на западном побережье Африки к югу до Нигерии, в Мали и Нигере (в последние, вероятно, попадает, пересекая Сахару), а также в странах восточного побережья Африки к югу даже до Кении и Танзании, где рост числа зимующих птиц связывают с ростом численности озерной чайки на территории бывшего СССР. Для чаек, гнездящихся на территории бывшего СССР, зимовки и миграции наиболее подробно изучены у восточнобалтийской популяции, объединяющей птиц Эстонии, Латвии, Литвы и Калининградской обл.. Эти птицы зимой встречены на обширной территории от Канарских островов и северо-западных берегов Африки до восточного побережья Черного моря (отдельные встречи — на Багамских островах и на Каспии), однако наиболее характерными местами зимовки этой популяции следует считать западное побережье Балтийского моря — юг Швеции, Дания, север восточной Германии, побережье Северного моря, включающее северо-запад западной Германии, Нидерланды, Бельгию, северный берег Франции и южную половину Британских островов, атлантическое побережье Франции и Пиренейский полуостров, озера и реки внутри континента (в основном в Германии, Швейцарии, Венгрии), а также западное Средиземноморье, особенно северное побережье Адриатического моря, долину р. По, юг Франции. Указанные зимовки достигаются в основном двумя путями: 1) чайки пересекают Балтийское море в западном направлении и попадают в южную Швецию и Данию, откуда достигают североморских и атлантических побережий Европы и Африки, а пересекая материк, попадают в Швейцарию и в западную часть Средиземного моря; 2) птицы двигаются вдоль восточного берега Балтийского моря, проходят через Калининградскую обл. (летевшие по первому пути в это время уже в Дании), попадают в Польщу и Германию, где пересекают материк и достигают Адриатического моря. Чайки, гнездящиеся в северной части Прибалтийского региона, более склонны к первому из описанных выше путей, гнездящиеся в южной части — ко второму. Чайки центральных областей европейской части России — Московской, Ивановской, Рязанской и Ярославской — зимой встречаются от юга Франции на западе до Каспия на востоке, однако их основными зимовками являются побережье Черного и юго-восточное побережье Азовского морей, побережья Италии, Югославии, Греции, восточное Средиземноморье (устье Нила, Ливан, Кипр и др.), среднее и верхнее течение Дуная, Швейцария. В ходе миграции к местам зимовок эти чайки примерно на 3 месяца задерживаются в среднем и нижнем течении Днепра, в низовьях Дона и на Азовском море, что дало основание выделить у них так называемый промежуточный перелет. Озерные чайки Казахстана и Западной Сибири зимуют на Каспии и предположительно также на берегах Персидского залива и Аравийского моря. Кольцеванием установлена зимовка камчатских озерных чаек в Японии. Миграции. Оставление гнездовых колоний начинается сразу после подъема молодых птиц на крыло, по срокам оно широко варьирует в зависимости от широты и местных условий в данном году и происходит в основном с третьей декады июня до начала августа. Озерной чайке свойственны послегнездовые кочевки, которые у птиц, гнездящихся вдали от границы ареала, имеют ненаправленный характер, а вблизи границы ареала могут иметь выраженную направленность и связаны с расширением ареала. Наиболее отдаленных зимовок озерные чайки достигают в середине декабря, в феврале их число там сокращается. На ближайших зимовках (например, Адриатика, юго-запад Балтики) чайки пребывают примерно с конца октября — начала ноября до середины марта. Сроки прилета сильно варьируют в зависимости от хода весны, но прилетает озерная чайка, как правило, несколько раньше освобождения водоемов ото льда. Например, в Эстонии в 1948—1966 гг. первые озерные чайки прилетали в среднем 7 апреля, наиболее ранняя встреча 23.III; в Калининской обл. за 36 лет наблюдений 26.III—23.IV, в среднем 7—8.IV, массовый пролет 1—17.IV; на оз. Киёво (в настоящее время колония прекратила свое существование) первые особи 24.III—8.IV, массовое появление 30.III—27.IV; в Барабинской степи первые особи в первой декаде апреля, пролет до двадцатых чисел мая, наиболее интенсивный — в конце апреля — начале мая. На Байкале (южная часть) озерная чайка появляется в середине апреля. В нижнем течении Сырдарьи первые птицы появляются в конце февраля, пролет в основном в марте, в Алма-Ате 10—19.III, под Семипалатинском 7—17.IV. Начавшиеся уже в XIX в. рост численности и расширение ареала в Западной Европе продолжаются во многих странах и в настоящее время. В 1970-х годах численность в странах Европы, кроме СССР, оценивалась минимум в 1400 тыс. гнездящихся пар. Более поздние публикации по отдельным странам дают основание предположить, что в этой части ареала гнездится от 1515 тыс. до 1820 тыс. пар озерных чаек. В том числе в Финляндии гнездится 150 тыс., в Швеции — 270 тыс., Дании — 210 тыс., Германии - 157 тыс., Польше - 84,5 тыс., Чехословакии - 200-350 тыс., Венгрии - 12 тыс., Нидерландах - 200 тыс., на Британских островах— 150-300 тыс. пар.Определение общей численности озерной чайки в бывшем СССР проведено для ряда западных территорий. В 1970-х годах она составляла: для Эстонии 80 тыс., Латвии 97 тыс., Литвы 30 тыс. пар, Белоруссии 104 тыс., Московской обл. 30—32 тыс. пар. Прослеживается и рост численности на упомянутых территориях: в Эстонии к 1960 г. гнездилось 20 тыс. пар, в 1967-1969 гг. — 30 тыс. пар, в Латвии в конце 1930-х - начале 1940-х годов — 10 тыс. пар, в 1964-1966 гг. — 30 тыс. пар, в Литве в начале 1970-х годов гнездилось 15-18 тыс. пар. Рост численности озерной чайки в последние десятилетия имел место также в других областях на северо-западе и в средней полосе европейской части России, а также вблизи населенных пунктов на Камчатке. Местообитание. В гнездовое время преимущественно мелководные внутренние водоемы — озера, пруды, водохранилища, дельты рек и т.д. — со сплавинами из корневищ надводной растительности или луговыми островами, торфяные карьеры, болота (преимущественно низинные), заросшие надводной растительностью морские бухты и морские острова вблизи берега. Реже гнездится на прибрежных лугах, дюнах, топких прибрежных кочкарниках и т.п. Местами добычи корма служат преимущественно открытые ландшафты - пахотные земли, луга и пастбища или водоемы. С усилением синантропизации местами кормежки все чаще становятся свалки, зверофермы, рыбоконсервные заводы и другие предприятия пищевой промышленности, рыболовецкие гавани, города. В период миграции больше связана с морским побережьем и поймами крупных рек, в период зимовки — с морским побережьем, сельскохозяйственными угодьями и населенными пунктами вблизи водоемов. Размножение. Начинает размножаться в возрасте 1—4 лет (т.е. после первой—четвертой зимовки), при этом самки начинают гнездиться в возрасте 1, 2 (в основном) и 3 лет, самцы — в возрасте 2, 3 (в основном) и 4 лет. Участие годовалых самцов в размножении, по-видимому, очень редкое . Моногам, диффузно-гнездящийся факультативно-колониальный вид. Колонии весьма постоянны и могут существовать в течение многих десятилетий, их перемещения вызваны обычно сукцессионными изменениями, продолжительным воздействием какого-либо неблагоприятного фактора (например, присутствие хищного млекопитающего), экстремальными условиями (например, затопление прежнего места колоний). По крайней мере часть чаек возвращается с зимовок парами, у большинства пары образуются после прилета вне колонии или в ней, но, как правило, не на месте будущего гнезда. Многие чайки меняют несколько партнеров, прежде чем приступят к постройке гнезда. До постройки гнезда, а также после вылупления птенцов чайки нередко меняют положение своего участка в колонии и переселяются в другое место. Первые прилетевшие чайки занимают будущий центр колонии, затем в течение нескольких дней занимается вся площадь, но с малой плотностью. С прибытием новых птиц и увеличением плотности наблюдаются центростремительные тенденции — чайки стараются переселиться в более предпочитаемые области колонии.Поведение озерной чайки изучено весьма подробно. Антагонистической демонстрацией в воздухе можно считать полет преследования и "налет и планирование": резкое увеличение скорости полета преследователя приводит к сходной реакции преследуемого, преследующая птица издает долгий крик и вибрирующий крик, после чего внезапно взмывает вверх, планируя на поднятых и отведенных назад крыльях, ноги при этом опущены, шея и голова вытянута вперед и вниз. Среди антагонистических демонстраций на земле следует выделить долгий крик в наклонной позе, являющийся предупреждением другой птице, приближающейся по воздуху, земле и воде. При этом голову птица держит поднятой высоко вверх и вперед, клюв более или менее горизонтально, красный зев хорошо заметен во время крика, запястные суставы отодвинуты от туловища; во время демонстрации птица обычно смотрит в сторону противника. Распластанная поза демонстрируется на воде и на суше . Агрессивная прямая поза — голова поднята вверх, оперение на шее и голове приподнято, клюв опущен, крылья немного приподняты. В этой позе птица либо стоит, либо двигается в сторону противника. Свидетельством готовности к бегству может служить испуганная прямая поза — шея также вытянута вверх, но оперение в ее верхней части прижато, клюв направлен более или менее горизонтально, запястные суставы немного приподняты, птица смотрит в сторону от оппонента.В территориальных спорах часто можно видеть демонстрацию мяукающего крика, которая обычно наблюдается после драки или атаки. "Квохтанье" особенно характерно в затянувшихся территориальных спорах (оно достоверно чаще наблюдается между соседями, чем между владельцем территории и чужаком), его часто можно видеть в плотных колониях. К антагонистическим демонстрациям относится также клевание земли. Отворачивание головы, по-видимому, служит позой умиротворения. Брачное поведение включает многие антагонистические демонстрации. При встрече самки самец издает долгий крик в наклонной позе, затем следует распластанная поза, которая отличается от агрессивного варианта приподнятым вверх клювом при опущенной горизонтально, на уровне туловища, шее (кроме того, в неагрессивных распластанных позах птицы обычно стоят бок о бок). В церемонию приветствия входит также отворачивание головы. Формирование пары завершается ритуальным кормлением. Самка выпрашивает корм в сгорбленной позе, издавая своеобразный звук.
Гнездовые колонии варьируют по величине от нескольких и нескольких десятков пар (в основном вблизи границ ареала) до многих тысяч (в средней полосе в условиях культурного ландшафта, где чайки освоили антропогенные источники кормов). Такие крупные колонии имеются на водохранилищах на Днепре (до 10 тыс), на озерах Бабите и Энгуре в Латвии. Самая крупная колония в Западной Европе находится в Нидерландах, в районе устья р. Шельда — 21 тыс. пар в 1976 г.. В условиях Прибалтики из крупных поселений (тысячи пар), занимающих несколько десятков квадратных километров, около 36% молодых птиц расселяются на расстояние до 500 км; самки с самцы одинаково часто. В пределах таких поселений (оз. Энгуре, 35 км 2) 45,6% оставшихся гнездятся на расстоянии 1,5 км от места рождения. Самцы более часто гнездятся в родной колонии (40,8%), чем самки (24,3%). В условиях оз. Энгуре около 20% выживших взрослых меняют гнездовую колонию; повторные кладки (бывают у птиц с возраста 3 лет) после отлова отмечались всегда в другой колонии, до 5 км от прежнего места. В колониях озерной чайки охотно селятся поганки (особенно черношейная, серощекая и красношейная), ряд уток (особенно хохлатая чернеть, также красноголовый нырок, серая утка, кряква и др.), речная крачка, малая чайка, пестроносая крачка и другие виды.Озерная чайка охраняет территорию 32—47 см в диаметре, оптимальное расстояние между гнездами примерно вдвое больше. Меньшее расстояние между гнездами бывает в случае, если что-нибудь препятствует визуальным контактам между насиживающими птицами. В Латвии на оз. Энгуре на 79 пробных площадках в 1974—1984 гг. плотность гнезд составляла 1,4—13,1 гнезд на 10 м 2, в среднем 5,3 гнезд на 10 км2; в радиусе 1 м 25,5% гнезд соседей не имело, 25,5% имело 1; 21,3% — 2; 14,7% — 3,..., 0,3% — 7 . На оз. Киёво к 1980 г. плотность колебалась в разных участках от 3—5 до 67 гнезд на 100 м2. На Украине в крупных колониях (1500—10000 пар) плотность достигает 2—5 гнезд на 1 м2, в мелких (100—250 пар) 8—10 гнезд на 100 м2. На Байкале в дельте Селенги в колониях величиной 16—1034 пар плотность гнезд составляла 0,5—7,2 на 100 м2, на Северном Байкале в колониях величиной 5—107 пар — 0,3—27,5 пар на 100 м2. В Северном Казахстане плотность достигает 32 гнезд на 100м2.Гнездо отличается от гнезд других чаек, населяющих сходные места, использованием более грубых материалов и некоторой небрежностью. Его размеры сильно варьируют в зависимости от расположения: например, на оз. Киёво гнезда на сухих и мокрых участках имели соответственно диаметр у основания 22 и 50 см, а высоту 4,5 и 30 см; в мокрых местах гнезда могут иметь еще большие размеры — диаметр 90 см, высоту 40 см.Сроки размножения варьируют на одних и тех же водоемах по годам в зависимости от хода весны, особенно уровня воды, поэтому выборочно приведем лишь самые общие сведения. В Латвии на оз. Энгуре первые яйца появляются в последней декаде апреля, средняя дата вылупления в 1974—1978 гг. на пробных площадках колебалась от 30.V до 9.VI, в целом по озеру на несколько дней раньше. В Литве в 1966—1973 гг. на оз. Жувинтас начало строительства гнезд отмечено 9—28.IV, первые птенцы появились 15—30.V. На водохранилище — охладителе, отличающемся повышенной температурой, первое яйцо в половине отложенных кладок в среднем появляется до 29.IV, т.е. на две недели раньше, чем в целом по республике. На оз. Киёво в 1933—1936 гг. первые яйца отмечены 17—24.IV, первые птенцы появились 18—23.V, начало массового вылупления в сухих рогозовых участках 22.V—2.VI; в 1975, 1977 и 1978 гг. на озере первые птенцы вылупились соответственно 3.V, 9—10.V, 13.V. Под Казанью первые яйца в 1974 г. отмечены 30.IV, массовая кладка 5—14. V. В Северном Казахстане первые яйца найдены 8—24.IV, первые птенцы — 16—18.V. На Байкале в дельте Селенги в 1973-1975 гг. в разных колониях начало яйцекладки зарегистрировано 7—13.V, массовая кладка проходила в период с 8 по 20.V в течение 3—8 дней, начало вылупления птенцов 1-12.VI; на Северном Байкале в 1972—1976 гг. начало яйцекладки отмечено 19.V—22.V, первые птенцы появляются 7.VI, обычно 15—18.VI. Более старые особи гнездятся раньше, чем трехлетние и двухлетние; при задержке начала размножения паводком различия в сроках сокращаются.В кладке 1-3 (чаще всего 3) яйца, возможность снесения одной самкой кладки из 4 яиц не доказана. Встречаются совместные кладки двух и нескольких самок, содержащие до 14 яиц; подобные кладки чаще всего наблюдаются при дефиците мест гнездования при высоком паводке. Средняя величина кладки за 5 лет в Латвии составляла 2,47—2,84 (обследовано 261— 415 кладок в год), в Литве — 2,68 ± 0,03 яиц; в ходе сезона наблюдается уменьшение среднего размера кладки. В Северном Казахстане средний размер кладки 2,7 (обследовано 2140 яиц). На Байкале в дельте Селенги в 1973-1975 гг. в разных колониях из 14-822 гнезд средний размер кладки составлял 2,03-2,70 яиц, на Северном Байкале в 1973-1976 гг. в отдельных колониях из 13-91 гнезд — 2,43-2,90 яиц. Если первое и последующие яйца вынимаются из гнезда в течение 12 ч после откладки, одна самка может снести в гнездо до 7 яиц. В размерах яиц не прослеживается географических вариаций. На оз. Киёво длина яиц составляла 50,54 ± 0,09 мм (п = 1348), ширина - 35,32 ± 0,07 мм (п = 1211), под Ижевском - 54,2 ± 0,17 Х 35,9 ± 0,12 (п = 175), на Северном Байкале в двух колониях 52,2 ± 0,36 Х 36,9 ± 0,4 (п = 52) и 51,9±0,18 Х 36,6 ± 0,15 (п = 60). Яйца сильно варьируют по окраске; чаще всего их основной фон буровато- или оливково-зеленоватый со сравнительно равномерно распределенными пятнами неправильной формы сероватого цвета (глубокие) и разных оттенков бурого (поверхностные). Наряду с нормально окрашенными встречаются также очень светлые голубоватые яйца, почти полностью лишенные рисунка.В насиживании участвуют оба пола, однако данные о их относительной роли противоречивы. По одним данным, по крайней мере в светлое время суток, в среднем больше насиживает самец (53,5%, наблюдения за 9 гнездами, всего 669 ч). По другим данным гораздо больше насиживает самка, при этом во время яйцекладки, ночью и в конце периода инкубации ее участие в насиживании особенно велико. Время от снесения до вылупления для 1-го яйца, по данным трех исследований, составляло соответственно 24,88 ± 0,81, 24,81 ± 0,07, 24,2 ±0,15 сут, для 2-го яйца — 23,59 ± 0,80, 23,87 ± 0,06 и 23,1 ±0,14 сут, для 3-го яйца - 22,82 ± 0,86, 22,73 ± 0,05 и 22,4± 0,16 сут.От появления первых трещин на яйце до вылупления птенца проходит 2—6, в среднем 3,3 дня. Последовательность вылупления в большинстве случаев соответствует последовательности снесения яиц. Обычно между вылуплением первого и второго птенца проходит 12 ч, между вылуплением второго и третьего — 24 ч. После обсыхания птенца скорлупа яйца удаляется; взрослая птица выбрасывает ее, отлетев на несколько метров. Масса только что вылупившихся и однодневных птенцов на оз. Энгуре в 1978 и 1979 гг. соответственно составляла 26,1 г (п = 510) и 25,4 г (п = 582). Масса птенцов в день вылупления на оз. Киёво (п = 85) 26,3 г, на севере Казахстана (п = 43) 24,8 г. В выкармливании птенцов участвуют оба родителя, по данным Т.И. Водолажской (1979), по крайней мере первые 5 дней жизни более активно птенцов кормит самка. В среднем птенцов кормят 5,7 раза в день, по другим данным — 4—5 раз, что составляет 100—150 г корма. Родители и птенцы узнают друг друга как визуально, так и по голосу через 3—4 дня. В первый день жизни птенцы обычно не проявляют тенденции к бегству, с 2—4 дня при опасности все чаще покидают гнездо, после 6-го дня бегство в определенные убежища — участки густой растительности — становится нормой; с возрастом длительность пробежек увеличивается. В убежищах птенцы разных выводков не проявлют взаимной враждебности, но их путь обратно к своему гнезду приводит к нападениям взрослых птиц — владельцев территорий и подросших (пятидневных и старше) птенцов. Примерно с 25—30-дневного возраста молодые ничинают летать, однако у части птенцов подъем на крыло затягивается на 2—3 недели. Как правило, взрослые оставляют колонию раньше молодых, самец часто раньше самки; по другим наблюдениям, родители нередко продолжают кормить и летных птенцов, часть взрослых отлетает из гнездовых колоний вместе со своими птенцами, и их контакт некоторое время сохраняется.Размеры гибели яиц и птенцов варьируют в широких пределах. Например, в Латвии на оз. Энгуре в 1974—1981 гг. процент вылупления сосатавлял 70,7—86,0% (1,75-2,34 птенца на пару), выжило до 25 дня жизни 37,2—66,4% из числа вылупившихся или 41,1—57,1% из числа снесенных яиц (0 ,94-1,55 птенца на пару). Смертность птенцов до 25 дня жизни в среднем составляла 42,9% (в отдельные годы 33,6—54,3%). В среднем 60,5% птенцов погибало до 4-дневного возраста, около 80% — до 10-го дня жизни. Новый пик смертности имеет место в момент перехода птенцов к самостоятельной жизни. На водохранилище Литовской ГРЭС за 3 года успешность вылупления составляла 81,8%, в среднем 2,19 птенца на кладку. В Белоруссии на Минском море за два года отход яиц составил 12,1%, постэмбриональная смертность - 57,5%, выросли в среднем 1,12 молодых на пару взрослых. На Байкале в дельте Селенги и на севере этого озера в нормальных условиях отход яиц составлял соответственно 20,0—32,5% и до 16% кладок, при наводнениях могут погибнуть свыше 90% кладок.Отмечены различия в успешности размножения в зависимости от сроков гнездования — наивысшие показатели соответствуют либо самым ранним, либо средним срокам. Погодные условия оказывают относительно меньшее влияние на смертность птенцов до 10-го дня жизни, чем на более старших птенцов, требующих больше корма. На первом году жизни после подъема на крыло смертность составляет ориентировочно 38%, взрослых особей 15% в год. Считается, что на рост численности озерной чайки большее влияние оказало сокращение смертности взрослых птиц, чем изменение успешности размножения. Максимальный возраст, по данным кольцевания, 32 года 1 месяц. Суточная активность, поведение. Активна как днем, так и ночью, в том числе и вне периода размножения; мигрирует также не только днем, но и ночью. Имеется два дневных пика активности — утро и ранний вечер. Общественна круглый год. В Западной Европе в местах ночевок зимой скапливается до 100 тыс. особей. Питание. Употребляет весьма разнообразные, преимущественно животные корма, легко переключается на наиболее доступные виды. В странах Западной и Средней Европы в период гнездования доминируют дождевые черви (до 50% общей массы) и насекомые (около 15%); летом и осенью значение дождевых червей уменьшается, чайки переходят на другие виды беспозвоночных, рыбу , фрукты и. семена растений, пищевые отбросы; на зимовках'насекомые поедаются редко, доминирует рыба и пищевые отбросы. Вследствие большого разнообразия условий в пределах ареала озерной чайки от этой общей схемы имеется много отклонений, которые в значительной степени определяются все расширяющейся и углубляющейся синантропизацией вида в значительной части ареала.Анализы содержимого желудков, собранных в мае—сентябре 1947—1959 гг. в Латвии показали доминирующую роль беспозвоночных в рационе. В частности, дождевые черви доминировали в мае, стрекозы, жуки и двукрылые — в июне, июле, августе; в сентябре увеличилось количество потребляемой рыбы, однако в целом она составляла лишь 3% по встречаемости. В рационе птенцов озерной чайки оз. Энгуре (побережье Рижского залива) встречаемость явно антропогенных кормов (отбросы пищевых продуктов, зерна злаков) и рыбы (добытой преимущественно в местах погрузки) менялась следующим образом: 1959 г. — 0% и 89,5%, 1963 г. — 233% и 52,0%, 1971 г. — 52,1% и 35,2%, что иллюстрирует радикальное изменение питания чаек этого водоема. В центре европейской части России основательные исследования питания озерных чаек проведены в 1930-1936 гг. на оз. Киёво. В ранневесенний период в рационе чаек преобладали мышевидные грызуны (в основном серая полевка) — 62,3%, насекомые и другие беспозвоночные, связанные с водой, — 26,1% встречаемости. В период насиживания доминировали личинки жуков (41,9%), наземные насекомые (26,2%); дождевые черви (28,3%), личинки жуков, живущие в почве (34,9%), и наземные насекомые (18,5%) преобладали также в период выкармливания птенцов. Животные, вредные для сельского хозяйства (майские жуки, их личинки, личинки щелкунов, серая полевка и др.), в период гнездования составляли в рационе 62,2% по встречаемости, полезные животные — 18,2%. Во время послегнездовых кочевок в рационе доминировали рыба (45,2%), насекомые (14,5%) и щитни (30,7%), а на зимовках на Каспии и Черном море рыба, добытая преимущественно у заводов и причалов. В 1970-е годы состав кормов чаек киевской колонии сильно изменился, так как основным местом кормежки стала свалка бытовых отходов. Для ряда других колоний средней полосы европейской части большую роль по сравнению с киевской колонией в период гнездования в рационе играет рыба.Преобладание насекомых в рационе в период гнездования констатировано также в Новосибирской обл.; в Казахстане весной в значительном количестве поедает мышевидных грызунов, летом — разных степных насекомых, в том числе пруса, осенью увеличивается потребление рыбы. На Байкале озерная чайка питается исключительно животными кормами, основу которых составляют беспозвоночные. Весной 50,3% содержимого желудков по объему составляли гаммариды, 30% — насекомые, моллюски, 113% — рыба. Летом насекомые (поденки, стрекозы, ручейники, двукрылые и др.) составляли 94% объема, рыба — 3,9%. Корм озерные чайки добывают разнообразными способами: плавая и передвигаясь пешком, в полете — с поверхности воды, земли или растений; ловят в воздухе. Дальность полетов за кормом зависит от численности птиц в колонии (группе колоний) и количества корма в окрестности. Она колеблется от нескольких до 70 км; для большинства птиц в крупных колониях все же не превышает 40 км.Враги, неблагоприятные факторы. Основную опасность представляет довольно широко практиковавшийся в прошлом неконтролируемый сбор яиц, что иногда приводило к исчезновению даже крупных колоний, и всякая хозяйственная деятельность (выпас скота и пр.) в местах колоний в период гнездования. Естественные враги те же, что и у других водоплавающих. Особую опасность представляют млекопитающие — лисица, енотовидная собака, американская норка, домашняя собака, кабан и др., однако часто гнездовья для большинства из них недоступны. Из пернатых значительный вред озерной чайке причиняют ворона, серебристая чайка, ворон, камышевый лунь, локально — филин, ястреб-тетеревятник и др. При этом успех хищничества вороны и серебристой чайки значительно увеличивается при беспокойстве колонии человеком. Резкий подъем уровня воды (например, во время штормов, на водохранилищах ГЭС) может полностью уничтожить все кладки в колонии. Основная причина гибели птенцов — недостаток кормов, наступающий, как правило, при продолжительной дожаливой прохладной погоде. При беспокойстве колонии гибель птенцов может достигнуть серьезных масштабов в результате агрессивного поведения взрослых по отношению к чужим птенцам. В литературе приводятся сведения о повышенной гибели птенцов, вызванной пастереллезом, ботулизмом, сальмонеллезами, а также о разных гельминтозах. Хозяйственное значение, охрана. Как массовый вид имеет весьма ощутимое значение в разных отраслях. Играет положительную роль в охотничьем хозяйстве, так как в гнездовых колониях озерной чайки плотность утиных кладок и их сохранность выше, чем в прилегающих сходных угодьях вне колоний. Легкое переключение на наиболее массовые виды кормов и большой радиус разлета за кормом определяют активное участие озерной чайки в уничтожении и ограничении численности вредителей сельскохозяйственных культур — насекомых и полевок — при массовом их появлении. Доказана возможность целенаправленного привлечения чаек на кормежку в определенных местах путем выставления макетов, что открывает возможность использования чаек для подавления локальных вспышек численности вредителей. Имеет определенное значение как санитар, собирая кормовые отбросы в звероводческих фермах и т. д. В Дании разрешен в установленные сроки сбор яиц озерной чайки, в ряде стран она считается охотничьей птицей (в Дании, Германии, Бельгии, частично в Австрии). Наряду с положительной ролью необходимо отметить создаваемую большими скоплениями чаек опасность для авиации, а также локально и кратковременно проявляющееся отрицательное воздействие чаек на прудовое рыболовство (уничтожение молоди). Однако вред, причиняемый рыбному хозяйству, сильно преувеличен. При необходимости регуляции численности рекомендуется ограничение доступа к антропогенным кормам, изменение гидрологического режима для устранения возможностей гнездования. Основные мероприятия по охране — обеспечение покоя в гнездовых колониях в период размножения.
|