Трофические связи
черноголовой чайки в гнездовой период в
условиях Черноморского заповедника
Т. Б. АРДАМАЦКАЯ, В. И. ВАКАРЕНКО, А. А. ПЕТРУСЕНКО
326240, Херсонская обл., Голая Пристань, Днепровская ул., д. 1. Черноморский государственный заповедник АН СССР
В условиях заповедника черноголовая чайка гнездится на островах. В поисках корма взрослые птицы посещают разнообразнейшие биотопы, о чем свидетельствует наличие в пищевых пробах птенцов представителей не менее чем 10 группировок организмов. Большинство составляли степные элементы (46,1%). Это прямокрылые
Calliptamus barbarus Costa, Myrmeleotettix sp., Euchorthippus pulvinatus F.-W., Oedipoda caerulescens L., маврская черепашка; жужелицы Amara similata Gyll., Zabrus tenebrioides Gz., пластинчатоусые Amphicoma vulpes F., Anisoplia austriaca Hbst., A. deserticola F.-W., чернотелки Anatolica eremita Stev., Pedinus borysthenicus Rchdt., долгоносики Mesagroicus obscurus Bohm., муравьи Messor clivorum Ruzs., Cataglyphis aenescens Nyl., Formica cunicularia Forst., Polyergus rufescens Latr. и др. Довольно значительным числом представлены политопные (ряд видов прямокрылых, полужесткокрылых, пластинчатоусых, щелкунов, коровок), различных двукрылых, особенно среди звонцов (27,4%), и луговые (некоторые жужелицы, долгоносики, чешуекрылые, в частности луговой мотылек, двукрылые) формы. На долю представителей других группировок (соленоводных, пресноводных, лесных, галофильных, болотных, пойменнолесных и супралиторальных) в целом приходится всего лишь 1,2%. Отсюда можно сделать вывод, что птицы собирают корм главным образом в степных биотопах и агроценозах. Довольно охотно они посещают и поименнолуговые биотопы. С другой стороны, не исключена возможность попадания луговых элементов на плакорные участки поливных земель.Согласно визуальным наблюдениям, в предгнездовой период чайки пролетают за кормом до 115 км. В гнездовое время, особенно после появления птенцов, радиус полетов сокращается до 50— 60 км, часто и меньше — до 5—45 км. При этом, согласно литературным данным (Клименко, 1950), зона полетов охватывает площадь в 4000 км
2, а в исследуемый период — видимо, значительно больше (до 6000 км2). Вполне естественно, что такая территория включает значительное количество мозаично перемежающихся биотопов. Этим и объясняется присутствие различных экотопических элементов в пищевых пробах. Тем не менее в каждой отдельно взятой пробе чаще всего присутствуют представители одной-двух, реже трех и лишь изредка четырех и более экотопических группировок. По-видимому, взрослые птицы собирают один пищевой комок, как правило, из одного биотопа.Другим не менее важным фактором, определяющим способ добывания корма, является ярусная приуроченность объектов питания, отражающая особенности вертикального размещения организмов. По данному признаку четко выделяется девять их группировок, причем доминировали обитатели растительных ярусов, или фитобионты,— 65,2%. В довольно значительном количестве обнаружены беспозвоночные, обитающие как в растительных ярусах, так и в опаде, или стратофитобионты,— 22,9%. В
меньшем числе отмечены организмы, связанные с поверхностью почвы, или эпигеобионты,— 5,6%; с растительным спадом, или стратобионты,— 4,0%; активно прокладывающие ходы в почве, или роющие геобионты,— 1,6% и в незначительном — беспозвоночные, попеременно обитающие в почве и опаде, или геостратобионты,— 0,1%. Из сказанного следует, что основную часть компонентов корма птицы собирают с поверхности растений, а также с поверхности почвы, куда они, будучи потревоженными, часто падают. Что касается роющих геобионтов, геостратобионтов и стратобионтов, ведущих скрытый образ жизни, следует иметь в виду, что эти организмы часто оказываются на поверхности при проведении культивации и других сельхозработ. Кроме того, в пасмурную погоду они часто активны и днем. Из водных обитателей попадались нектонты, планктонты и бентонты, вместе составившие 0,6% что еще раз подтверждает известный вывод о том, что черноголовая чайка питается практически почти полностью на суше.В пространственном отношении трофические связи животных тесно связаны и с характером распределения популяций представителей отдельных их видов. Поэтому компоненты питания черноголовой чайки были условно разделены на две группы — образующие различные скопления в виде колоний, роев, при массовом выплоде и т. д. и равномерно-диффузно распределяющиеся по всей поверхности субстрата, с которой собирается корм. Доминирующее положение в пище птенцов исследуемого вида заняли представители первой группы — саранчевые
(Calliptamus italicus L., С. barbatus Costa), пластинчатоусые родов Gyrnnopleurus 111., Onthophagus Latr., Aphodius 111., Oniticellus Serv., имаго и гусеницы лугового мотылька, муравьи и двукрылые из сем. Chironomidae (всего 17 видов), вместе составивших 67,0% от общего количества обнаруженных пищевых объектов.К временным аспектам питания относится потребление животного корма разных онтогенетических стадий составляющих его компонентов, возраст потребителей, а также характер суточной активности тех и других. Нами обнаружено подавляющее доминирование взрослых (имагинальных) форм (75,5%). Личинки и куколки обнаружены в значительно меньшем количестве (9,6%), остальное приходится на беспозвоночных, у которых возрастные стадии определить трудно (малощетинковые черви, моллюски, ракообразные, паукообразные и др.— 11,6%) и растительные корма. Вероятно, это связано с преобладанием в корме подвижных компонентов (63,2%), большую часть которых как раз и составили имагинальные формы насекомых. Это большинство жуков-жужелиц, щелкуны, чернотелки, муравьи и др., среди которых сравнительно небольшим количеством представлены летающие формы (23,0%) — большинство прямокрылых, чешуекрылые, ряд видов перепончатокрылых, двукрылые. Довольно значительной была доля (36,9%) сравнительно малоподвижных и неподвижных объектов — малощетинковые черви, моллюски, пластинчатоусые жуки, листоеды, долгоносики и др. Очевидно, бегающие и летающие формы беспозвоночных чайкам трудно доступны, о чем свидетельствует их незначительная (4,6%) доля в рационе.
Для характеристики возрастных стадий потребителей мы располагаем пищевыми пробами, взятыми у птенцов трех возрастных групп: младшего (6—10 дней), среднего (11—20 дней) и старшего (21—35 дней). Количество наименований компонентов питания в этих группах колебалось от 51 до 128; причины таких колебаний не выявлены. Обнаружено только, что в 1974 г. в пищевых пробах птенцов старшего возраста было значительное количество растительных кормов, тогда как в 1975 и 1977 гг. они не были отмечены. Среди кормов животного происхождения в 1974 г. у птенцов старшего возраста преобладали прямокрылые, жуки-чернотелки, муравьи, в меньшем количестве попадались чешуекрылые и двукрылые. В 1975 г. количество муравьев, полужесткокрылых и жесткокрылых резко снизилось. Указанные обстоятельства требуют дополнительных исследований. Противоречивы данные и относительно наполненности пищевых проб. Так, среднее количество компонентов на одну пробу у птенцов младшего возраста в 1974 г. составляло 130 экз., среднего — 208, старшего — 70, тогда как в 1975 г.— 73, 60 и 287 соответственно.
Весьма неожиданным было подавляющее доминирование (50,3%) в собранных днем пробах компонентов с сумеречно-ночным типом активности, прежде всего массовых видов — лугового мотылька, звонцов и др., собираемых птицами с поверхности растений, где эти насекомые проводят дневное время суток. Другие беспозвоночные с подобным типом активности, как уже отмечалось, часто бывают доступны птицам при проведении сельхозработ. Беспозвоночные с дневной активностью в рационе птенцов составили 29,4%, а с круглосуточной — 20,3%.
Функциональная характеристика трофических связей
Собранный и обработанный материал позволяет включить в данный раздел отдельные обобщения, касающиеся способов поиска и распознавания объектов питания по отдельным морфоэкологическим признакам, а также роли исследуемых птиц в трофической сети населяемых ими экосистем. Относительно поиска и распознавания объектов питания трофические связи можно проследить через соотношение последних по ряду других морфоэкологических признаков, из которых выделены характер подвижности,
склеротизации, окраски и линейные размеры. Соотношение компонентов по первому признаку уже обсуждалось в связи с их сопоставлением по отдельным онтогенетическим стадиям. По степени склеротизации эти организмы условно разделены на объекты с покровами мягкой, средней и твердой консистенции. В пище птенцов преобладали первые (50,6%), две другие группы были представлены примерно в равном соотношении (соответственно 26,6 и 22,8%). Это вполне естественно, поскольку птенцам, особенно в раннем возрасте, необходимы более питательные и мягкие корма; среди таких кормов в пробах отмечались личинки пластинчатоусых, гусеницы чешуекрылых, звонцы и пр. По данным же В. Д. Сиохина, в пище взрослых птиц изучаемого вида в условиях Присивашья и Молочного лимана преобладали компоненты с твердыми покровами. По характеру окраски среди животных компонентов основную часть составили субстратные формы (63,9%) — прямокрылые, многие полужесткокрылые, ряд видов чешуекрылых, двукрылых и др. Это объясняется тем, что именно среди таких животных преобладали подвижные формы. Среди объектов с контрастной окраской преобладали малоподвижные и неподвижные формы — личинки и куколки чешуекрылых, двукрылых жесткокрылых и т. п.Спектр линейных размеров пищевых компонентов птенцов оказался довольно широким — от 2 мм (моллюск
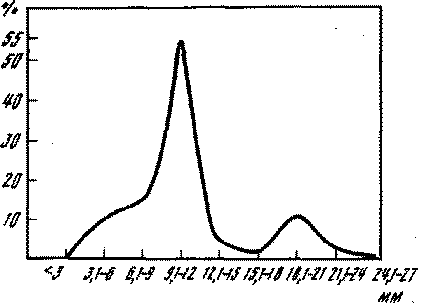
Bittium reticulatum Costa, муравей Tapinoma kirnburni К. Агп.) до 120 мм (кольчатая сколопендра Scolopendra cingulata L.). Из приведенного графика (рис. 1) видно, что преобладали компоненты размерами от 9,1 до 12,0 мм; их было 55%. На графике виден еще один небольшой пик (10,2%), соответствующий размерам 18,1—21,0 мм. Первый пик объясняется тем, что мезофауна исследуемых экосистем отличается доминированием компонентов именно этих размеров. Второй пик получился за счет отдельных видов саранчевых, жесткокрылых, а также гусениц лугового мотылька.Самым главным аспектом изучения значения того или иного вида в исследуемых экосистемах является прежде всего определение соотношения компонентов его питания по признаку трофической специализации. Безусловно, такие расчеты следует проводить в весовых показателях, но поскольку мы ими в достаточной мере не обладаем, то нам приходится оперировать количеством особей в выделенных трофических группировках. В корме черно
Рис. 1. Количественное распределение пищевых компонентов по линейным размерам
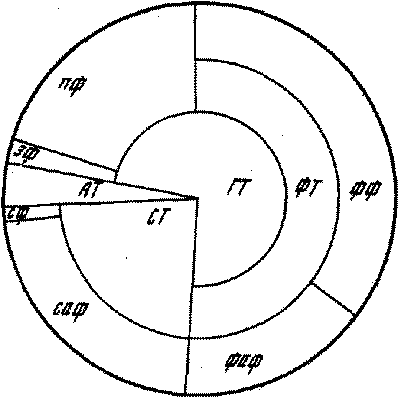
Рис. 2. Трофическое воздействие черноголовой чайки на биокомпоненты экосистем в гнездовой период
AT — автотрофы;
ГТ — гетеротрофы;
СТ — сапротрофы;
ФТ — фитотрофы;
ЗФ — зоофаги;
ПФ — пантофаги;
ФФ — фитофаги;
ФАФ — афаги фитотрофного происхождения;
СФ — сапрофаги;
САФ — афаги сапротрофного происхождения
головых чаек преобладали афаги (38,9%), т. е. онтогенетические фазы животных, практически не принимающие пищи либо полностью обходящиеся без нее. Это имаго лугового мотылька, звонцов, куколок насекомых с полным превращением и т. д. Второе место занимают фитофаги (35,7%), третье — пантофаги (20,6%). В незначительном количестве обнаружены зоофаги (1,0%) и сапрофаги (0,4%). Помимо этого, зарегистрированы рассмотренные выше растительные корма (3,4%). Если учесть, что указанные афаги на других стадиях своего развития потребляют либо растительную пищу (гусеницы лугового мотылька), либо являются сапрофагами (личинки звонцов), то картина воздействия черноголовой чайки в гнездовой период на компоненты населяющие его экосистемы значительно изменится (рис. 2). При этом основную часть составят фитофаги (51,1%), за ними будут следовать сапрофаги (23,8%) и пантофаги. Таким образом, исходя из анализа полученных данных, можно судить о том, что в гнездовой период черноголовая чайка в наибольшей степени воздействует на гетеротрофов (фитофаги, зоофаги, пантофаги — 72,7%), в меньшей степени — на сапротрофов и в незначительной степени — на автотрофов.
Практическое значение
Из всего списка объектов питания черноголовой чайки животного происхождения (295 наименований) около половины (150) известны как вредители и переносчики заболеваний культурных растений, и в количественном отношении они составили лишь немногим менее половины (45,0%) общего объема проанализированных проб. Из них основную часть (26,0%) составили чешуекрылые, главным образом за счет лугового мотылька (24,0%). Из других вредителей более или менее многочисленными оказались прямокрылые (4,2%), пластинчатоусые (3,5%) и долгоносики (1,0%). Из отдельных видов в большом количестве отмечены маврская черепашка (7,0%) жужелицы родов Amara Bon. et Zabrus Cirv. (5,5%), кузьки Anisoplia Latr. (2,7%).
Если учесть, что колония черноголовой чайки в Черноморском заповеднике насчитывает более 330 тыс. пар и добавить к ним примерно 9—25% холостых птиц, то число добытых ими вредителей превысит 100 млн особей за один день и 5 млрд за гнездовой период. Из этого количества более 1,5 млрд особей приходится на долю вредителей зерновых культур, среди
которых основное место занимают вредители пшеницы. Из обнаруженных в пищевых пробах вредителей к ним относятся хлебная и некоторые другие жужелицы, клопы-черепашки и кузьки; кроме того, черепашки и личинки хлебных жужелиц часто препятствуют колошению хлебов.Заключение
Данное сообщение не претендует на полное раскрытие рассматриваемого вопроса. Главная его задача заключается в поисках эффективных методик, направленных на возможно более детальное изучение экологии черноголовой чайки через анализ сопоставления потребляемых им объектов по выделенным качественно-количественным и морфоэкологическим признакам. В частности, обнаружено, что при весьма обширном спектре корма (около 300 названий), свидетельствующем о широкой экологической пластичности черноголовой чайки, более или менее обычными оказались компоненты всего 16 названий, а массовыми — только 8.
Обобщение материала по признакам ярусно-биотопической приуроченности объектов питания, с одной стороны, отразило мозаику биотопов региона, а с другой — преимущественный сбор корма из степных экосистем и агроценозов. Кроме того, оно фактически подтвердило результаты визуальных наблюдений, касающихся сбора пищи большей частью с поверхности растений, в меньшей степени — из растительного спада, поверхности почвы и в незначительной — из верхних слоев почвы и с водоемов. Ряд не менее интересных и уже приведенных выводов сделан при сопоставлении компонентов по другим признакам — характеру распределения популяций, численности, подвижности, онтогенетическим стадиям суточной активности, окраске и линейным размерам.
Соотношение объектов по пищевой специализации наглядно показало роль черноголовой чайки в трофической сети исследуемых экосистем. Прежде всего отмечено, что в гнездовой период данный вид наибольшее воздействие оказывает на представителей блока гетеротрофов, прежде всего на консументов первого порядка (фитофагов). В практическом отношении около половины его рациона составили вредители и переносчики заболеваний культурных растений.
ЛИТЕРАТУРА
Клименко М. И. Материалы по фауне птиц Черноморского заповедника // Тр. Черномор, гос. заповедника. 1950. № 1. С. 8—15.
SUMMARY
Trophic relations of the Mediterranean Gull during the nesting period at the Black Sea State Reserve. Report II. The spatial-temporal charasteristics of trophic relations. Ardamatskaya T.B.,Vakarenko V.I.,Petrusenko A. A.
Based on the long-term studies at the Black Sea State Reserve, the materials on the characters of the layer-biotopical confinement of food places, which had expressed the mosaic of the regional biotopes and the principal food from the steppe ecosystems and agrocenoses. Daring the nesting period near halt of the diet of the Mediterranean Gull contained pests and carriers of cultural plant deseases.